





探索蛋白质-蛋白质相互作用:对理解健康与疾病的关键作用
蛋白质-蛋白质相互作用(PPIs)的探索历程已逾四十载,其研究之初主要依赖于蛋白质亲和层析技术和抗体共免疫沉淀等手段逐步推进。随着科学技术的日新月异,PPIs的分析范围已实现了从单一相互作用向基因组层面的飞跃。这一进展得益于细胞内标记技术的革新、亲和纯化结合质谱(AP-MS)技术以及共分级质谱(CF-MS)方法的引入。
时至今日,科研人员已能够借助交联质谱(XL-MS)和冷冻电子显微镜(cryo-EM)等尖端技术,更为精准地辨识直接蛋白质相互作用与间接蛋白质相互作用之间的微妙差异。这些高精尖技术,加之人工智能工具(诸如AlphaFold等)的广泛运用,预示着PPIs及蛋白质复合物研究领域即将迎来前所未有的深刻洞察。这不仅将极大地丰富我们对基础生物学的认知,更为疾病机制的探究开辟了全新的视角,有望为基础生物学研究和疾病治疗策略的制定带来革命性的突破。

引言
生命的复杂性源于构成细胞干质量约90%的生物大分子,这些大分子在细胞中扮演着至关重要的角色。其中,蛋白质以其多功能性脱颖而出,不仅催化了细胞中绝大多数必要的化学反应,还发挥了不可或缺的结构性作用。这些功能的实现往往需要蛋白质相互结合,形成蛋白质复合物。许多这样的复合物以“蛋白质机器”的形式存在,通过有序的结构变化,为细胞提供有益的构象转变。部分蛋白质机器已被广泛研究,其组成成分能够在体外经过纯化后重新组装,并进行高分辨率的结构分析。例如,DNA转录和复制所涉及的多蛋白复合物就是两个典型的例子。
在探索这些蛋白质机器的过程中,科学家需要建立高度灵敏且特异性的检测方法,并结合遗传学分析,以识别并纯化出细胞中催化特定生化过程的所有组分,进而进行功能表征。这种方法不仅揭示了包括蛋白酶体、染色质重塑复合物、剪接体和核糖体在内的多种蛋白质机器的分子机制和结构特征(其中剪接体和核糖体的研究还突显了RNA在这些机器中的核心作用),也推动了我们对生命科学的深入理解。
然而,值得注意的是,即便是最简单的已知活细胞——一种经过基因工程改造的支原体,其473个对繁殖至关重要的基因中,仍有近三分之一的功能尚未明确。而对于人类基因组,尽管我们认识到一个基因可能具有多种功能,但在约20,000个蛋白编码基因中,仍有数千个基因的功能未知。这不禁让人思考,维持生命可能还需要多少尚未被发现的活性?实际上,我们的细胞中可能隐藏着数百种尚未被表征的蛋白质机器,而传统的生化检测方法却难以触及它们。
理解这些未知的蛋白质机器对于揭示基本生物学过程和疾病状态背后的生物学机制至关重要,而这些知识正是开发新型治疗手段所急需的。因此,蛋白质-蛋白质相互作用(PPI)图谱的绘制为解决这些问题提供了一个充满希望的途径。通过这一途径,我们有望揭示更多未知的蛋白质机器,进一步推动生命科学的发展,并为疾病的诊断和治疗提供新的思路。
利用蛋白质亲和层析技术深入洞察蛋白质-蛋白质相互作用(PPIs)对生物功能的影响
随着科学技术的持续进步,细胞蛋白质-蛋白质相互作用(PPIs)的研究领域也迎来了同步的发展。鉴于蛋白质与其他分子间的相互作用通常展现出高度的特异性,早期的PPIs识别尝试便聚焦于蛋白质亲和层析技术。自20世纪初以来,该技术便被广泛用于纯化与特定分子结合的蛋白质,或用于阐述酶及其他蛋白质与其底物、辅因子等相互作用分子的关系。
在随后的岁月里,亲和层析方法在支撑材料的选择和配体偶联技术上取得了诸多改进。特别是,不溶性抗原被用于从血清中纯化特异性抗体,这一应用极大地推动了亲和层析技术的发展。此外,琼脂糖珠的引入以及氰基溴(CNBr)活化琼脂糖的方法,使得蛋白质及其他分子能够方便地通过一级胺基偶联到琼脂糖上,从而显著提升了实验的效率和灵活性。
尽管理论上可以通过固定化的纯化蛋白在细胞提取物中无偏倚地寻找未知的相互作用伙伴,但这一方法直至20世纪80年代才首次被应用于寻找与噬菌体λ和T4蛋白相互作用的蛋白质。例如,以λ N蛋白(一种转录抗终止因子)作为配体的亲和层析实验,成功地从大肠杆菌蛋白中筛选出了一个与琼脂糖珠支持物无非特异性结合的蛋白,即NusA。进一步的研究揭示,NusA是λ N蛋白与RNA聚合酶相互作用的适配因子。同时,其他宿主辅助因子,如NusB、核糖体蛋白S10和NusG,也通过亲和层析技术被证实与转录抗终止调控系统的其他组分存在相互作用。
这些研究充分展示了蛋白质亲和层析技术在检测弱PPIs方面的强大能力,其能够检测到亲和力低于10⁵ M⁻¹的相互作用。此外,其他方法,如融合蛋白的高浓度拉下实验、在细胞中过表达带标签的蛋白、或交联蛋白后进行亲和纯化等,也展现出了在检测弱相互作用方面的潜力。这些技术的不断发展,为我们更深入地理解PPIs对生物功能的影响提供了有力的工具。

蛋白质亲和层析在早期面临两大主要技术挑战。首先,该技术需要高纯度的蛋白质作为配体。然而,在20世纪80年代,通过传统的柱层析方法纯化蛋白质往往既耗时又复杂,开发针对特定蛋白质的纯化方案可能需要耗费数年时间。此外,尽管当时已经引入了重组DNA技术(大约在20世纪70年代中期),但可供克隆的基因数量仍然有限,蛋白质过表达载体尚未得到广泛应用,同时,用于简化蛋白质纯化过程的亲和纯化标签也尚未被引入。
其次,识别通过亲和层析捕获的相互作用蛋白也是一个难题。如果研究的生物系统中的蛋白质组分已经被纯化,并且它们在SDS-聚丙烯酰胺凝胶上的迁移率已知,那么识别这些蛋白质就相对容易。然而,对于那些尚未通过遗传学方法鉴定的相互作用蛋白,研究者就需要依靠推测其功能,并通过一系列的生化实验来验证这些推测。这种方法虽然既耗时又费力,但在20世纪80年代确实促成了一些重要发现,例如TFIIF亚基的纯化与鉴定,以及大量果蝇肌动蛋白丝和微管结合蛋白的识别。这些成就证明了即使在面临技术局限的情况下,科学家们依然能够通过创新和毅力取得突破。
通过抗体和融合蛋白识别PPIs的生物学新视角
在蛋白质相互作用研究的历程中,抗体和融合蛋白成为了蛋白质亲和层析的重要替代手段。早期实验中,研究者利用针对特定纯化蛋白的抗体,通过免疫沉淀技术从细胞提取物中成功捕获了目标蛋白及其相互作用伙伴。例如,未知的53 kDa宿主蛋白(后确认为肿瘤抑制因子p53)与SV40大T抗原的结合,以及肿瘤抑制蛋白RB与其他病毒致癌蛋白的相互作用,都是通过这种方法得以揭示。此外,针对病毒致癌蛋白pp60v-src及其细胞同源物pp60c-src的单克隆抗体,能够共免疫沉淀出含有磷酸酪氨酸的蛋白,这一发现强调了SH2结构域与磷酸酪氨酸肽段间直接相互作用在细胞信号传导中的核心作用。
另一项关键进展是利用针对果蝇TATA盒结合蛋白(TBP)的单克隆抗体,成功共免疫沉淀了RNA聚合酶II通用起始因子TFIID的其他亚基(即TBP相关因子TAFs),这不仅鉴定了这些亚基,还推动了其分子克隆和在不同物种中的功能表征,确立了它们作为转录共激活因子的角色。
随着蛋白克隆技术的普及,设计融合蛋白以连接不同合作伙伴与目标蛋白成为新的研究策略。谷胱甘肽-S-转移酶(GST)和麦芽糖结合蛋白(MBP)等融合伙伴的引入,使得通过亲和层析一步即可纯化目标蛋白。此后,多种蛋白标签如金黄色葡萄球菌蛋白A、多组氨酸序列和小肽标签(如HA标签)等被广泛应用,极大地丰富了蛋白纯化手段。
在数以千计的实验中,融合蛋白被固定在琼脂糖珠上,并与细胞提取物混合以“拉下”相互作用蛋白。例如,在癌症生物学研究中,重组SH2结构域与大肠杆菌TrpE蛋白的融合成功拉下了酪氨酸磷酸化的表皮生长因子(EGF)受体和血小板衍生生长因子(PDGF)受体。
免疫共沉淀与拉下实验的局限性
尽管这些方法为生物学研究提供了宝贵信息,但免疫共沉淀在扩展到多蛋白研究时面临挑战。高纯度目标蛋白和高特异性抗体的制备往往困难重重,且抗体可能干扰蛋白复合物的组装。同样,拉下实验也可能因融合伙伴阻碍目标蛋白与相互作用伙伴的结合或复合物组装而失败。此外,在基因组测序时代之前,克隆编码目标蛋白的基因或cDNA既耗时又繁琐,且过表达的融合蛋白在异源系统中可能难以保持可溶性或正确的翻译后修饰。
AP-MS:通过细胞内标记、亲和纯化与质谱识别PPIs
质谱技术的引入为相互作用蛋白的识别带来了革命性变化。在基因组测序完成前,偶尔可通过Edman降解法结合cDNA克隆和测序的机会识别某些相互作用蛋白。然而,电喷雾电离(ESI)和基质辅助激光解吸电离(MALDI)等软离子解吸方法的开发,使质谱技术能够测定包括肽和蛋白在内的大分子的质荷比(m/z)。这一突破推动了胰蛋白酶等氨基酸特异性蛋白酶消化后肽段质量的MS测定,并与DNA或氨基酸序列数据库比对,从而识别目标蛋白。液相色谱-串联质谱(LC-MS/MS)的进一步开发,极大地提高了复杂混合物中肽段的识别能力,为大规模蛋白质组学研究奠定了坚实基础。
综上所述,抗体和融合蛋白在PPIs识别中发挥了关键作用,但免疫共沉淀和拉下实验存在局限性。随着质谱技术的引入和发展,特别是AP-MS技术的应用,为蛋白质相互作用研究开辟了新的视野。

细胞内蛋白标记与亲和纯化结合质谱(AP-MS)的生物学研究
通过在天然宿主细胞中表达带有特定标签的蛋白,随后利用与标签结合的固定化试剂纯化目标蛋白及其相互作用蛋白,以供质谱(MS)分析,这一方法展现了诸多优势。它避免了异源系统中过表达蛋白可能引发的不溶性问题,确保了带标签蛋白能够获得天然的翻译后修饰(PTMs),这些修饰对于蛋白的溶解性及其与其他蛋白的结合至关重要。此外,在天然伴侣蛋白和辅伴侣蛋白的辅助下,目标蛋白能够正确折叠。通过将标签DNA整合到编码目标蛋白的基因中,标签蛋白能够以接近天然的水平表达,从而减少了因过表达而导致的假性相互作用的风险。
AP-MS的生物学见解
在酿酒酵母和大肠杆菌等模式生物中,细胞内染色体蛋白标记和串联亲和纯化(TAP标签)技术的运用,使得对几乎所有蛋白进行AP-MS研究成为可能。这种系统化的基因组范围蛋白质-蛋白质相互作用(PPIs)和蛋白复合物鉴定,为生物学研究提供了许多重要见解。例如,它揭示了染色质生物学中的多个关键机制,包括COMPASS复合物介导的组蛋白甲基化、SET3复合物介导的组蛋白去乙酰化,以及Rpd3组蛋白去乙酰酶在启动子和转录区域中的特定功能。此外,还发现了能够用组蛋白H2A的变体Htz1替代原有组蛋白的特化染色质重塑复合物。
在多细胞动物中,AP-MS技术结合现代质谱和生物信息学方法,成功鉴定了目标蛋白及其广泛的相互作用蛋白网络。以果蝇和人类细胞系为例,研究生成了包含多达5000种蛋白的PPIs网络。这些研究不仅深化了对疾病相关信号通路的理解,还揭示了多种蛋白修饰在调节PPIs中的关键作用,如Hippo信号通路中STRIPAK激酶复合物介导的磷酸化依赖相互作用。
AP-MS的局限性
尽管AP-MS技术功能强大,但其也面临一些挑战。首先,标记蛋白的过表达有时可能导致伪相互作用的出现,尽管利用CRISPR技术进行细胞内染色体标记可以在一定程度上避免这一问题,但目前该方法的应用仍局限于少量目标蛋白。其次,某些生理上相关但较弱的相互作用可能在AP清洗步骤中解离,从而无法被检测到。此外,使用全细胞提取物的方法都存在一个共同问题,即某些蛋白在体外可能发生相互作用,但在体内由于定位或表达时序的差异,实际上并不具备相互作用的能力。对于膜蛋白等难溶性蛋白的识别,更是增加了挑战,尽管目前使用温和的去污剂可以在一定程度上解决这一问题。
共分级-质谱(CF-MS)表征蛋白质复合物的生物学见解
CF-MS技术基于一个早期蛋白质纯化研究中的原则:如果多肽在多步纯化过程中共同分离,则它们可能属于同一蛋白质复合物。CF-MS通过色谱分级后对蛋白质进行胰蛋白酶消化,并利用串联质谱(MS/MS)进行识别和定量。该方法无需对宿主或病原体的所有蛋白进行标记或纯化,即可分析其相互作用,特别适合研究病原体感染的细胞。同时,CF-MS能够揭示病原体对宿主PPI网络及蛋白复合物的间接影响。然而,由于共纯化的蛋白可能是偶然结合,因此CF-MS识别的PPIs和蛋白复合物需要与已知数据进行对比验证。
PPIs的验证及区分直接和间接相互作用
对于通过高通量方法(如AP-MS或CF-MS)识别的PPIs,在进一步研究或开发治疗方法之前,需要进行严格的验证。可信的PPIs通常表现为在遗传研究中属于同一生物学系统、定位于同一细胞内区室,或在不同细胞类型和干扰条件下表达量高度相关。为了验证PPIs并区分直接和间接相互作用,研究者们开发了多种方法,包括免疫沉淀-免疫印迹法(IP-Westerns)、FRET和BRET、近距离连接检测(PLA)以及生物素标记技术(如BioID和TurboID)等。然而,这些方法通常难以完全区分直接和间接PPIs。因此,制备重组蛋白并在体外测试相互作用,以及利用遗传方法(如酵母双杂交系统,Y2H)识别二元PPIs,也是常用的验证手段。
交联-质谱(XL-MS)区分直接PPIs
交联-质谱技术通过使用双功能交联试剂处理分离的蛋白质复合物,并进行LC-MS/MS分析,可以可靠地识别复合物中邻近的肽段。这种方法不仅可以验证体内发生的PPIs,还能识别在AP过程中因清洗步骤而丢失的弱PPIs。此外,XL-MS能够明确介导相互作用的蛋白表面,为开发干扰PPIs的药物提供重要信息。
利用AP-MS研究野生型和突变型人类及病毒蛋白的疾病见解
近年来,研究PPIs的时间和空间变化成为前沿课题。了解PPIs如何在细胞的不同区域中改变,以及在环境条件变化时随时间发生的动态变化,对于解析细胞功能至关重要。AP-MS技术已被广泛应用于探索与疾病相关的特定突变对PPIs的影响,包括神经退行性疾病、癌症、自闭症谱系障碍(ASD)以及病毒感染等领域。这些研究不仅揭示了PPIs的生物学连通性和收敛性,还将基因发现置于蛋白复合物和功能通路的框架下,从而更有效地捕捉生物学上的共性和机制。
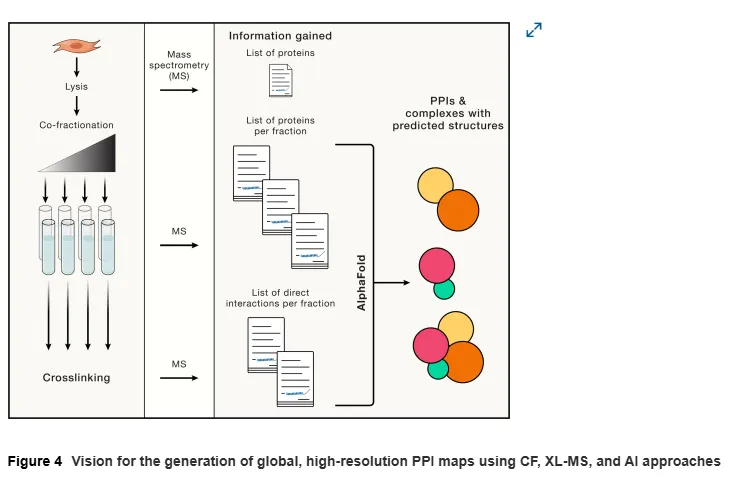
综上所述,通过结合CF-MS、AP-MS和XL-MS等多种技术,研究者能够更全面地理解蛋白质复合物的组成和功能,为基础生物学研究和疾病治疗提供新的视角和策略。

蛋白质机器与复合物的生化解析及功能理解
随着科技的不断进步和资源的持续积累,科学家们已经能够利用亲和纯化(AP)技术分离出所有稳定组装的蛋白质复合物,并通过质谱(MS)技术对其蛋白组成进行鉴定。尽管蛋白质复合物的亚基组成和功能可能因细胞类型的不同而有所差异,但共分级-MS(CF-MS)技术的出现,使得我们能够揭示特定细胞类型中复合物的亚基组成,从而确定最适合纯化每种复合物版本的细胞类型。
然而,蛋白质复合物在细胞中往往以多种形式存在,即使在同一细胞中,也可能存在具有不同但部分重叠亚基组成的复合物版本。例如,在酿酒酵母中,Rpd3催化亚基就参与了两种不同的组蛋白去乙酰酶复合物:Rpd3L复合物和Rpd3S复合物,它们分别在不同的转录调控过程中发挥作用。这些复合物可以通过传统的生化分级方法或在特异性亚基上添加AP标签进行解析,这表明互补的生化方法在解析包含重叠蛋白的复合物组成方面具有重要作用。
类似地,人类SWI/SNF染色质重塑复合物(BAF复合物)也存在多种版本,这些版本在癌症和神经发育障碍中尤为常见。通过生化和结构方法对这些不同版本的BAF复合物进行区分,为理解其功能提供了重要线索。
蛋白质复合物功能的优先研究方向
在获取蛋白质复合物列表的基础上,解析这些复合物的功能成为更为关键的一步。当数据不足时,科学家们可以优先研究那些被怀疑为“蛋白质机器”的复合物。蛋白质机器通常由多个基因产物组成,其功能依赖于能量驱动的单向构象变化,能够充当分子马达、时钟或组装因子等角色。这些机器的动力源通常是高能键的水解,如ATP或GTP,因此保守的核苷酸结合位点可以作为识别可能具有机器特性的复合物的标志。
研究蛋白质机器功能的方法
解析蛋白质机器功能的标准方法通常涉及纯化组分并重建系统。通过结合使用蛋白标签来简化蛋白质的纯化过程,科学家们可以利用动力学和化学分析来研究其不同组分的反应。此外,蛋白质机器的多构象转换特性使其成为冷冻电子显微镜(cryo-EM)技术的重要研究对象。如今,通过亲和纯化将感兴趣的复合物直接捕获到冷冻电镜栅格上,已经极大地简化了实验过程。最新的cryo-EM技术还能够从混合物中识别复合物的“构象景观”,进一步提高对新型蛋白质机器的理解。
AI与Cryo-EM结合AP-MS和CF-MS在PPI靶向疗法开发中的应用
在解读差异性蛋白质-蛋白质相互作用(PPI)数据集中的突变效应时,了解哪些突变通过直接影响特定界面来扰动PPI至关重要。然而,AP-MS数据的一个主要局限在于难以区分直接PPIs与通过其他蛋白介导的间接PPIs。虽然交叉链接质谱(XL-MS)和Cryo-EM能够解决这一问题,但前者缺乏全面性,后者操作繁琐。
人工智能(AI)技术的最新进展正在改变这一局面。例如,AlphaFold通过高精度预测蛋白质结构,已经能够成功预测PPIs。这些预测有助于筛选直接相互作用并优先研究与疾病相关的突变。基于这些预测结果,科学家们可以将直接PPI界面的突变优先用于基因研究和基于结构的Cryo-EM分析。
在将PPI数据应用于治疗方面,PPI数据可以帮助识别新的治疗靶点并揭示如何“靶向”这些靶点。例如,通过分析头颈癌相关的PIK3CA突变与HER3的相互作用,科学家们发现抑制HER3功能可能是一种有效的治疗策略。而如果突变破坏了PPI,则通过分子胶恢复该相互作用可能具有潜在价值。此外,PPI数据还可以用于开发双功能蛋白降解诱导剂(PROTACs)和分子胶降解剂等新型药物。AI技术还可以帮助预测与PPI界面特异性作用的化合物,进一步推动精准治疗的发展。
结论
过去,研究者们曾试图仅从疾病相关的基因组测序数据中直接提炼出生物学和治疗学的深刻见解。然而,实践证明,仅凭序列信息所能揭示的奥秘相当有限,且随着对特定疾病领域进行更深入的测序研究,所取得的进展往往呈现出收益递减的趋势。事实上,细胞的功能核心在于蛋白质,它们通过物理相互作用实现“沟通”,进而组装成复杂的分子机器,这些相互作用几乎涵盖了细胞的所有生理功能。更为关键的是,与疾病密切相关的突变往往会干扰蛋白质的正常功能及其相互作用,而当前大多数用于治疗疾病的药物也是直接针对蛋白质设计的。因此,对蛋白质-蛋白质相互作用(PPIs)进行深入且机制化的理解,对于揭示健康生物学的奥秘以及探索疾病治疗的新途径具有举足轻重的意义。
尽管PPI研究的历史已近半个世纪,但近年来实验技术与计算方法的整合取得了突破性进展,为我们以高分辨率的方式识别和表征稳态PPIs以及受疾病突变影响的PPIs提供了前所未有的机遇。这些由数据驱动的整合策略为我们开启了全新的视角,标志着PPI研究迈入了一个崭新的时代。这一新纪元不仅将推动我们对健康与疾病的分子生物学机制进行更深入的剖析,还将为开发针对多种疾病的新型治疗方法奠定坚实的基础。
| 名称 | 货号 | 规格 |
| PIST, CT, Control Peptide (PDZ Protein Interacting Specifically with TC10, CFTR-associated ligand, CAL, dJ94G16.2, Fused in Glioblastoma, FIG, Golgi-associated PDZ and Coiled-coil Motif Containing, Golgi Associated PDZ and Coiled Coil Motif Containin | P4207-51A-50ug | 50ug |
| PDZ Domain Containing 3 (PDZD3) Antibody (FITC) | abx308382-1mg | 1mg |
| PDZ And LIM Domain 2 (PDLIM2) Antibody | abx431633-200ul | 200ul |
| PDZD2 (PDZ Domain-containing Protein 2, Activated in Prostate Cancer Protein, PDZ Domain-containing Protein 3, AIPC, KIAA0300, PDZK3) (MaxLight 650) d | P3129-81B-ML650-100ul | 100ul |



 危险品化学品经营许可证(不带存储) 许可证编号:沪(杨)应急管危经许[2022]202944(QY)
危险品化学品经营许可证(不带存储) 许可证编号:沪(杨)应急管危经许[2022]202944(QY)  营业执照(三证合一)
营业执照(三证合一)